- Research Paper
- Open access
- Published:
Climate and ungulate browsing impair regeneration dynamics in spruce-fir-beech forests in the French Alps
Annals of Forest Science volume 79, Article number: 11 (2022)
Abstract
Key message
Different components of water balance and temperature reduce density and height growth of saplings of Picea abies (L.) H. Karst (Norway spruce), Abies alba Mill. (silver fir) and Fagus sylvatica L. (European beech) in mixed uneven-aged forests in the French Alps and Jura mountains. Ungulate browsing is an additional pressure on fir and beech that could jeopardise the renewal of these species in the future.
Context
The uncertainty in tree recruitment rates raises questions about the factors affecting regeneration processes in forests. Factors such as climate, light, competition and ungulate browsing pressure may play an important role in determining regeneration, forest structures and thus future forest composition.
Aims
The objective of this study was to quantify sapling densities and height increments of spruce, fir and beech and to identify dominant environmental variables influencing them in mixed uneven-aged forests in the French Alps and Jura mountains.
Methods
Sapling height increment and density were recorded in 152 plots, and non-linear mixed models were obtained to establish relations between them and environmental factors known to affect regeneration, namely altitude, slope, aspect, canopy openness, soil characteristics, temperature, precipitation and ungulate browsing.
Results
Regeneration density, varying from 0 to 7 saplings per m 2, decreased with sapling height and was also negatively affected for spruce by PET, but positively for fir by precipitation and for beech by mean annual soil water content. Height increment reached up to 50 cm annually, increasing with sapling height and canopy openness and decreasing under high maximum summer temperatures for spruce and beech. The statistical effect of different environmental variables varied slightly among species but trends were quite similar. Additionally, ungulate browsing was high, with fir being the most intensely browsed, followed closely by beech, while spruce was rarely browsed.
Conclusions
All these results suggest that more temperature warming and a decrease in water availability could negatively impact sapling growth and density in the three species, with possible reduction of forest renewal fluxes. The observed increase of ungulate populations leading to increased browsing could be particularly detrimental to fir saplings.
1 Introduction
Mixed uneven-aged forests of Norway spruce (hereafter spruce), silver fir (hereafter fir) and European beech (hereafter beech) occupy extensive areas in Europe (Brus et al. 2011), specially in the French Alps (Toïgo et al. 2015). Their presence throughout Europe, in such wide environmental conditions, makes them a target of rapidly changing climatic extremes such as increasing frequencies and intensities of droughts (Cailleret et al. 2013; Diaci et al. 2020) and ever increasing summer temperatures (Schurman et al. 2019; Lombaerde et al. 2020). The last few decades have also witnessed increasing number of wild ungulates in the Alps, which also puts plants in these forests at a higher risk of being browsed (Heuze et al. 2005; Nagel et al. 2015; Bernard et al. 2017). Different studies have highlighted changes in plant demography (Gray and Brady 2016), inconsistent demographic responses to climate at species range edges (Kunstler et al. 2020), limitation of regeneration and tree recruitment (Silva et al. 2012; Dey et al. 2018; Löf et al. 2019), stressing a potential risk of reduced forest regeneration. For a species to maintain its range, regeneration must compensate mortality and harvesting in a climate warming context. Several articles document and predict increasing frequency of mortality events because of changing climate (Bodin and Wiman 2007; Allen et al. 2010; Manso et al. 2015). Natural regeneration in many forests is especially variable and already low (Dey et al. 2018). If regeneration reduces while mortality increases with climate change, even if harvesting events remain constant, this could jeopardise forest renewal. A diminution of regeneration because of global changes could therefore have a major impact on species distributions and their persistence.
In mountain forests, regeneration processes are especially slow and scarce. Regeneration has often been stressed as a limiting process in mountain forest dynamics (Kräuchi et al. 2000). To better understand and anticipate degraded regeneration dynamics and potential species resilience or adaptability, it is necessary to identify key drivers of regeneration demographic processes. These include processes such as seed production, germination, and seedling growth, mortality and survival, up to the stage of tree recruitment. Our objective in this study was to quantify sapling densities and height growths of spruce, fir and beech in mountain forests and to identify key environmental factors influencing them. Studies such as Balandier et al. (2006); Diaci et al. (2020) have shown the role of many different factors in these processes. Stands of spruce, fir and beech are known to largely depend on light availability for their regeneration growths (Dyderski et al. 2018; Diaci et al. 2020; Lombaerde et al. 2020; Gaudio et al. 2017). The role of soil nutrients and water availability in facilitating regeneration is also well studied (Madsen and Larsen 1997; Dyderski et al. 2018). Besides, regeneration depends on a large scale on inter- and intra-specific competitors. However, the species identity and cover determine if plants can act as facilitators or competition to certain species (Balandier et al. 2006; Collet and Chenost 2006; Laurent et al. 2017; Thrippleton et al. 2017). Disturbances such as fires (Dey et al. 2018), frost (Defossez et al. 2015) and insect infestations (Dobrovolny 2016) also affect regeneration processes. In spite of this knowledge, accurate estimation of regeneration demographic rates and precise identification of the effects of climate and other environmental factors on regeneration in mountain forests is difficult. This could be because saplings are scattered in space with high variability among sites, environmental factors could be highly inter-correlated, and saplings are difficult to monitor over successive years because of their small size and low survival rates. We tried to overpass some of these difficulties by analysing sapling densities and height increments by sampling a large environmental gradient in the French Alps and Jura mountains.
In this study, we address the following questions related to the regeneration of spruce, fir and beech in mixed uneven-aged forests:
-
Studies have observed and predicted high variability as well as decreasing regeneration densities and height increments. What are the magnitudes of sapling height increments and densities in the French Alps and Jura mountains?
-
Many factors such as light, temperature, soil conditions, water availability and presence of ungulates are known to affect regeneration. What are the key drivers of sapling height increments and densities of spruce, fir and beech?
We also discuss if there is a risk to the current species distribution and forest structures because of regeneration shortage and unreliability.
2 Materials and methods
2.1 Field sampling
2.1.1 Study sites and plot sampling strategies
The study was carried out in the French Alps (44.17∘ N, 5.23∘ E to 46.39∘ N, 6.90∘ E) in mixed uneven-aged mountain forest stands, with at least one out of spruce (Picea abies (L.) H. Karst), fir (Abies alba Mill.) and beech (Fagus sylvatica L.) as dominant species. Sampled plots were distributed over 11 mountain ranges, to cover a wide range of climates, elevations and site conditions (Fig. 1, Table 1). The sampling locations were determined based on existing networks of permanent plots established by the Office National des Forêts (ONF), France and INRAE, with information of the species tree basal area on the plots. The plots in these networks were first categorised into 24 classes, and then subsets taken to cover various stand structures and environmental conditions in the sampling. The categorisation was based on the following variables:
-
Total basal area (BA) of adult trees on the plot (2 classes: higher and lower than the mean of all plots)
Fig. 1 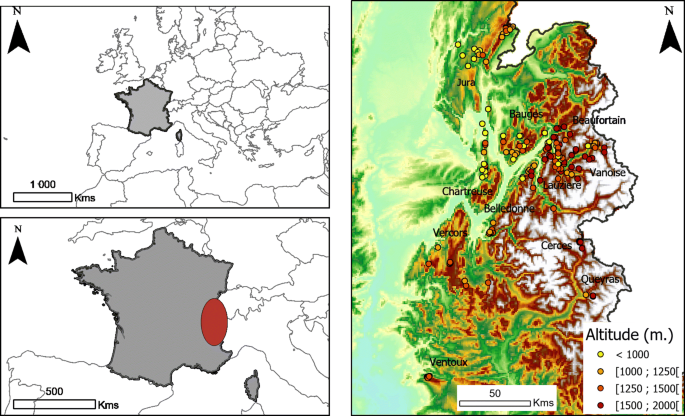
Locations of sampled plots and mountain ranges in the French Alps and Jura mountains, with each point representing a sampled plot, and colours of points representing altitude gradients
Table 1 Site-wise distribution of sampled plots, with distributions of altitude, temperature and plot basal area covered -
Mean annual air temperature (T∘C) of the plot (2 classes: higher and lower than the mean of all plots)
-
Annual mean of monthly precipitation (mm) on the plot (2 classes: higher and lower than the mean of all plots)
-
Identity of the dominant species on the plot (spruce, fir or beech)

Thus, we covered a wide range of stand structures, tree densities and compositions (BA of the different species as a proxy) for the different climates encountered in the area, including the distribution edges of the species (temperature and precipitation as a proxy). Thus, 24 categories were formed, and up to five plots were chosen by random selection from each category. A total of 152 plots were sampled across all mountain ranges. The observations described in the following sections were carried out by the authors. All data is available here: Unkule et al. (2021).
2.1.2 Assessing stand structure
In order to characterise stand structure and composition, two concentric circular plots of radii 10 m and 15 m were marked from the centre, to measure the tree basal area (BA) for each species. Diameter at breast height (DBH) of all adult trees above 7.5 cm and 17.5 diameter were measured in the 10-m and 15-m radius circle, respectively, using a tape or a caliper, and the species identity noted. This information was used to calculate tree BA per hectare and per species for the plot.
2.1.3 Assessing sapling growth, density and browsing rates
The Point Center Quadrat Method (PCQM) was used to estimate sapling density, with a maximum radius of 10 m (Mitchell 2015). This method was selected after a survey of different regeneration sapling methods (Carnet et al. 2018). A big advantage of this method is that it makes it possible to adjust the prospected area to sampling density and avoids empty plots due to small plot sizes.
The 10-m radius circular plot was divided into four quadrants, with boundaries in the north, east, south and west directions. Saplings were divided into 4 height classes: 10–50 cm (H1), 51–150 cm (H2), 151–300 cm (H3) and above 300 cm, but below DBH of 7.5 cm (H4). This division was made in order to evaluate, if any, an effect of the sapling height. Across 152 plots, we sampled 2370 individual saplings (plants between height 10 cm, and DBH <7.5cm), 720 spruce, 891 fir and 759 beech.
In each quadrant (i), the sapling nearest to the plot centre, of each species, and in each height class, was selected, and the following measures noted to the smallest count of 0.5 cm: sapling length when held straight (H); mean of annual shoot length increments for the past 3 consecutive years (ΔH), at least when it was possible; and distance (Di) (in m) of sapling from the centre of the plot (Fig. 2). The annual shoot increments were determined by measuring the length between marks of two bud scars, which are known to be indicators of one growing season. Using this information, the density of saplings of each species and height class was computed as: \((1/((\sum D_{i})/(N_{0}/N_{Q}))^{2}) \times CF\) where N0 is the total number of quadrants (here 4) and NQ is the number of vacant quadrants. CF is a correction factor obtained from a table developed by Warde and Petranka (1981) to take into account the presence of vacant quadrants. As the role of wild ungulates on sapling growth and survival is often highlighted, any physical damage to the terminal bud of a sapling was noted as an incident of browsing by an ungulate, in order to quantify the browsing rate for each plant species. The height increment of browsed saplings was considered zero.

Representation of a PCQM plot used for estimating sapling densities: each dot represents the sapling nearest to the plot centre in each quadrat, for each height class and species measured (maximum radius = 10m)
2.1.4 Calculation of competition and browsing pressure indices
A competition index was calculated to quantify competition of spruce, fir or beech with other understory vegetation (all plants less than 150 cm height, either herbaceous or woody species). First, dominant understory plant species were identified. Their cover was estimated by a visual assessment of ground cover occupied by the vertical projection of their foliage onto the soil (GC). The different species were assigned scores (SC) according to their potentially negative competition effect on regenerated saplings. Very strong interfering species such as gramminoids and dense ferns were ranked 3, less competitive species such as Rubus sp. 2, tall shrubs and midstorey trees 1 and other small forbs 0 (SC) (Balandier et al. 2006). A competition index was computed for each quadrant as: \(\sum GC*SC\), which was then averaged for the plot. Additionally, an approximate percentage of browsed plants on the plot was noted (BP), and the browsing pressure index was computed as: \( (\sum GC*BP)/\sum GC\). These indices were calculated using approximate visual covers of the vegetation.
2.1.5 Evaluating canopy openness
Canopy openness (O) of a site influences stand micro-climate and light. In each quadrant, canopy openness was measured in the four cardinal directions at a distance of 4 m from the centre, using a convex densiometer (Baudry et al. 2013).
2.1.6 Laboratory analyses of soil samples
Within a 1-m radius around the plot centre, a soil sample was collected, up to 20-cm depth (excluding litter and humus) using a soil auger. This sample was later used in the laboratory to estimate the C:N ratio and pH of the soil.
Soil samples were first air dried in the lab and were then placed in aluminum trays in a ventilated oven at 40∘C. After complete drying, they were manually sieved through a 2-mm mesh (Retsch steel analytical sieve, ISO 3310-1 standard). Next step was to fine grind them with a 0.25-mm mesh for carbon and nitrogen determination (using an ultracentrifugal mill ZM200 Retsch, stainless steel rotor 8 teeth). Carbon and nitrogen were detected on the 0.25-mm sample by a total combustion process, using an elemental analyser by the company Vario Micro Cube, Elementar, whereas the pH determination was done on the 2-mm-sieved samples using a SevenExcellence pH meter from Metler-Toledo, InLab Solids Pro-ISM electrode.
2.1.7 Soil and climate data extracted from digital maps
To complement data from field surveys, soil and climate information was extracted, described during the period from 1985 to 2010 for each plot, using digital maps belonging to the Digitalis database by Laboratory SILVA, Université de Lorraine-AgroParisTech-INRA (https://silvae.agroparistech.fr/home/). Monthly solar radiation calculated using the Helios model (Piedallu and Gégout 2007), minimum and maximum temperatures modelled and mapped using Meteo France weather stations and potential evapotranspiration (PET) calculated with Turc formula (Turc 1961; Piedallu et al. 2016) were used to represent available energy.
Precipitation (Prec) and climatic water balance (CWB, Prec minus PET) were used to estimate available water for plants from climatic data. Soil water balance distribution was also estimated, combining monthly climatic variables with soil water holding capacity maps (SWHC) using the Thornthwaite formula (Thornthwaite 1955) with a Geographical Information System. This calculation provided the spatial distribution of mean annual values of actual evapotranspiration (AET), soil water content (SWC) and soil water deficit (SWD) (Piedallu et al. 2013).
The resolutions of all the digital maps was 50 m, except precipitation and SWHC, which had 1-km resolution. All climatic data were provided as monthly average of the 1985–2010 period, which were later also aggregated into seasonal and annual means.
2.2 Data analyses
All data analyses and models were calibrated separately for the three species, spruce, fir and beech, using R software version 3.6.0 and version 4.0.2 (R Core Team 2017), and figures were produced using the package ggplot2 (Wickham 2009). Two different datasets were used for the analyses of densities and growth of saplings. For the density analyses, plot-level data were used, with sapling densities for each species and height class pooled at the plot scale (n = 152). For the analyses of growth of saplings, each data point consisted of information of a single individual sapling (n = 2370). The final models determining annual height increment was formulated using the subset of unbrowsed saplings (n = 2128), as the height increment of browsed saplings was assumed to be zero.
2.2.1 Analysis strategy
The variables potentially affecting sapling density and growths were grouped into categories described in Section 2.1: sapling height, climate, soil, topography, competition and stand structure (Table 2). A systematic approach was used to formulate multivariate non-linear mixed models, testing the progressive addition of variables from a new category. Preliminary exploration indicated that the variable influencing sapling densities and height increments the most was sapling height. We tested then the addition of stand structure variables such as canopy openness, total and conspecific tree basal area. In the next step, we added climate and soil category variables to the earlier model, such as maximum summer temperature, minimum winter temperature, mean monthly precipitation, annual mean of monthly soil water content, soil water deficit, soil water holding capacity, its CN and pH. The last step was to test the addition of competition (competition index, browsing index), followed by topographical factors (slope, altitude, aspect). The order of these steps was chosen based on the importance of these variables derived from literature, and considering increasing pressure of climate change. When a variable from one category was selected, other variables from the same category were systematically avoided to prevent correlated variables being present in the same model (except topographical factors). The function “nlme” from the R package “nlme” (Pinheiro et al. 2021) was used to run these models. Evaluation of the models was done by comparing graphical predictions made by different models and their ecological significance, and by comparing the observed vs. predicted values, along with the Akaike Information Criteria (AICs), mean square deviations (MSD) and the distribution of residuals. A p-value of 0.05 was deemed significant. Based on the principle of parsimony, the simplest model with the most accurate predictions was selected. Even though a number of variables were tested, the selected models were relatively simple, with 2 predictive variables for sapling densities and 3 for height increments.
2.2.2 Sapling densities
The model that explained sapling densities (Nijk) for height class i of species j on plot k, as a function of sapling height (Hijk) and a climate variable (Vjk), when significant, different for each species was:
aj and cj were species-dependent parameters and bjk were parameters depending both on species and plot.
2.2.3 Unbrowsed sapling height increments
The other equation was inspired by the formulation of height increment of conifers by Ligot et al. (2020). A similar approach as earlier was followed for obtaining models explaining annual height increments. The chosen model for sapling height increments (ΔHijk) for an individual i of species j on plot k, as a function of sapling height (Hijk), canopy openness (Ok) and a climate variable (Vjk), different for each species, was:
Both equations had a random plot effect, which was tested on different parameters.
2.2.4 Browsing
Due to the absence of height increment on a browsed sapling, it was not possible to include it as a factor in the earlier models. Hence, a preliminary exploration was carried out to identify the extent of browsing pressure and its possible damage. Browsing proportions of each species, and saplings of different heights were compared using 2 proportion z-tests, in order to estimate similarities and differences between different proportions. A proportion test (R function “prop.test” (Newcombe 1998)) was used to make comparisons, by specifying the alternative hypothesis to find out if the browsed proportions of each category are significantly different from one another.
3 Results
3.1 Main factors affecting sapling densities
Out of 152 sampled plots, 116 plots (76%) displayed some spruce regeneration, 128 fir regeneration (84%) and 104 beech regeneration (68%). However, the most obvious observation was the overall low regeneration densities (Table 3). Globally, even though only 3 plots (1.9%) displayed no regeneration, 40 plots (26%) had total sapling densities below 0.1 sapling per m2, and only 22 plots (14.4%) displayed total sapling densities above 1 sapling per m2. Only 2 plots had total sapling density of more than 3 saplings per m2, indicating the low levels of regeneration.
Even though survival rates were not recorded, it was observed that smaller saplings of all three species (<50 cm height) were much more abundant than taller saplings (Fig. 3). Density of saplings of height above 50 cm quickly tended towards zero, which suggested the possibility of survival of very few saplings to older ages.

Simulation of sapling regeneration densities along sapling height gradients of spruce (a), fir (b) and beech (c) in mixed uneven-aged mountain forests. For each species, colored lines represent the sapling density changes with different values of the climate or soil variables selected by the models (PET mean monthly potential evapotranspiration (mm), Precannual mean of monthly precipitation (mm), SWC annual mean of monthly soil water content (mm)). Values of the variables correspond to first quantile, median and last quantile of the observed values. Warning: Scales differ on the y-axis
The equations and parameters representing models that best explained sapling densities are shown in Table 4. The graphic representations shown in Fig. 3 represent predictions and confidence intervals (95%) from these models. All the models that were tested are described in Table 5.
The selected models showed that densities of saplings in all three species decreased exponentially with increasing height. Additionally, spruce densities decreased exponentially with increasing mean monthly PET (Fig. 3a); fir densities increased exponentially with increasing mean monthly precipitation over the year (Fig. 3b); and beech densities increased exponentially with increasing annual mean of monthly soil water content (SWC) (Fig. 3c). Apart from that, a random effect was observed for sapling densities, which could be a result of many other potential plot-specific factors that could not be separated and/or quantified. This effect was stronger for spruce, followed by beech, and least for fir (Fig. 3).
Among all climate and soil variables (V) tested in Eq. 1, even though the most significant ones were selected for each species, there were some other models that were significant as well, but did not make sense ecologically. For example, the density model for beech with height and PET had the lowest AIC value, but the model produced negative sapling densities, and had to be discarded. Additionally, some significant models had higher AIC values (Table 5). For example, spruce sapling densities were more affected by temperature and precipitation variables, but not so much by soil water conditions. Fir and beech however showed more sensitivity to soil nutrients and soil water conditions. Unfortunately, addition of biotic factors like the competition index and browsing index to the model was not found to be significant. The level of significance of these models also differed, as described in Table 5.
3.2 Main factors affecting unbrowsed sapling height increment
The measured annual height increment of saplings varied from 0 to 50 cm. The increment of unbrowsed saplings was highest for beech, followed by spruce, and fir displayed the lowest annual height increment, specially for saplings taller than 300 cm. The same approach and environmental variables were tested for models explaining height increments, as for sapling densities.
The most significant model showed that the annual height increment of an unbrowsed sapling depended on species, height, canopy openness and the maximum summer temperature observed on the plot (for spruce and beech). The equations and parameters representing models that best explained sapling height increments are shown in Table 6, whereas the results shown in Fig. 4 represent predictions and confidence intervals (95%) from these models.

Prediction of annual sapling height increment of spruce (a), fir (b) and beech (c) along canopy openness (O) gradient, and for 3 sapling heights, 40, 150 and 500 cm, in mixed uneven-aged mountain forests. For each species, colored lines represent the height increment changes with different values of the best climate or soil variables selected by the models (Max Sum temp, maximum summer temperature (∘C)). Values of the variables correspond to first quantile, median and last quantile of the observed values
Among our sampled species and individuals, the annual height increment of an unbrowsed sapling was most importantly limited by the height of the sapling. Smaller saplings typically grew slower, with differences among species. Canopy openness (hereafter, O) also determined the growth levels of saplings. The results of the selected model (explained in Section 2.2) revealed that the growth of all saplings was very low at canopy openness below 10%, growing rapidly as it increased, following a power function and reaching almost a horizontal curve at canopy openness levels above 40%. Taller saplings reached this horizontal asymptote at higher canopy openness than smaller saplings.
Saplings belonging to different species also displayed different maximum growths. Beech saplings grew most rapidly followed by spruce, while the least height increment was recorded by fir.
Beech and spruce growth was negatively affected by maximum summer temperature (Fig. 4a, c). Silver fir growth did not directly depend on any other environmental factors that we took into account in our dataset (Fig. 4b). Apart from that, plot-specific differences were observed in sapling height increments as well, indicating a considerable random effect. This effect was stronger for beech, followed by spruce, and least for fir.
The significance of other tested climate and soil variables was less obvious for height increment of saplings of all species than for densities, based on high p-values for other variables (Table 7). The closest models strengthened the dependence of beech and spruce on temperature, as they included different temperature variables like minimum winter temperatures and maximum monthly temperatures.
3.3 Browsing rates
Out of a total of 2370 saplings that we documented, 242 saplings were browsed or damaged by ungulates at the terminal shoot. This number varied greatly among saplings of different heights and species. Among the different height classes, the most browsed were saplings below 150 cm, which were easily accessible to most browsers. However, within the three species, there was a clear difference in the preference of wild ungulates. As seen in Fig. 5, and confirmed by the two-proportion z-tests, fir was the most browsed species among the three (14%), followed by beech (11%). However, the z-test showed no significant difference between the total browsing proportions of the two species (p = 0.127). Spruce was the least browsed with 2% of all the saplings browsed, much less than beech (p = 3.447e −16) and fir (p = 1.128e −11).

Comparative browsing rates of saplings by species (ABI: fir, FAG: beech, PIC: spruce) and height classes (1 =10–50cm, 2 = 51–150cm, 3 = 151–300 cm, 4 = 301cm–DBH < 7.5cm), as well as mean browsing rates per species. Letters a, b, c and d represent statistically similar and different bars
4 Discussion
4.1 Understanding regeneration densities
As observed by forest managers and scientific studies such as Kräuchi et al. (2000); Lines et al. (2019), our study sites had low overall regeneration density of spruce, fir and beech. On an average, all three species had a large number of saplings lower than 50 cm height (Table 3), with a steep decrease in the number of taller saplings. This suggested that the reason for low total sapling densities was not the limitation of seed rain or germination, but low survival rates (Simon et al. 2019). As annual survival rates were not recorded, we relied on diminution of densities across height classes to evaluate survival. Fir densities decreased most rapidly with sapling height, indicating high early mortality, possibly due to high browsing rates of fir. This has also been observed by Nagel et al. (2015) and Bernard et al. (2017). There was no direct effect of browsing observed on sapling densities. However, through its strong effect on growth, browsing maintained saplings at lower heights, and higher mortalities, reducing the density of taller saplings indirectly. Comparison of sapling densities with enclosed experimental plots is necessary to clarify this effect of browsing on density (Bernard et al. 2017). We, however, identified other key factors affecting sapling densities and height increments.
As stand conditions are often the result of multiple historic events (climate extremes, management, browsing pressure etc.), it was impossible to disentangle the different effects, or put in evidence general rules of what is a compromising factor in a given environment (Benavides et al. 2016). Models were tested to evaluate the effect of light (Ligot et al. 2014; Orman et al. 2021; Lochhead and Comeau 2012), basal area (Stancioiu and O’Hara 2006), water availability (soil and atmospheric) (Löf and Welander 2000; Moser et al. 2016), temperature (Schurman et al. 2019; Lombaerde et al. 2020), competition with other species (Thrippleton et al. 2017), browsing (Bernard et al. 2017) and many other variables that are known to affect regeneration densities (Table 2). We succeeded in identifying the most dominant variables affecting spruce, fir and beech densities, among the ones that we tested. However, the extent and intensity of these effects was species specific (Caron et al. 2021). Spruce regeneration densities improved in lower PET (potential evapotranspiration) levels, and significantly declined in higher PET levels, which is supported by Cienciala et al. (1992), who demonstrated high sensitivity of spruce trees to evapotranspiration. Among all the models tested, there were sometimes more than one significant models. In the case of spruce, all significant models (that were not selected) indicated temperature sensitivity of spruce. None of the models with soil conditions as an explanatory variable (except C:N) showed any significance. This is in contradiction to Dyderski et al. (2018), who showed the importance of soil pH and water logging for spruce regeneration. Beech densities increased with higher SWC. Many studies support this result as soil water availability is known to have an effect on both beech saplings and adults (Gessler et al. 2006; Pröll et al. 2015; Diaci et al. 2020). Fir densities relied least on climate factors, though it showed some sensitivity to annual precipitation, requiring higher water input to survive better. Though there are studies that indicate that fir can survive soil water deficits (Tinner et al. 2013), some also predict that the species will most likely suffer in severe drought conditions (Bottero et al. 2021). However, it is interesting to note the detrimental effect of water constraints on the three species, with either limitations in water input, uptake or evaporation. A look at Fig. 3 also suggests that the effects are stronger for spruce and beech and that they are more climate sensitive than fir. There are multiple studies discussing tree and sapling drought tolerance of the three species which conclude that all three species show signs of drought sensitivity (Cailleret et al. 2013; Löf et al. 2005; Silva et al. 2012), though there is a debate about their comparative sensitivities (Diaci et al. 2020; Schall et al. 2012; Zang et al. 2014; Vitali et al. 2017).
We assumed fecundity (or seed rain) of a species to be directly proportional to its conspecific basal area on the stand (similar to Paluch et al. (2019)), and we expected sapling densities be directly proportional to the conspecific BA as well. However, higher BA also reduces light availability, which can hinder regeneration. There was no clear relation between regeneration and BA, perhaps because the two effects could compensate each other, or maybe our plot size was too small for BA to be a good predictor of the local seed rain, or BA was not a good predictor of light at this local scale (the canopy is too heterogeneous). Similar observations have been made by Comeau et al. (2006).
There was high variability in the regeneration data collected both within and between plots. The variability within a plot was probably due to the strong heterogeneity of the environmental factors experienced by individual saplings (Schurman et al. 2019; Diaci et al. 2020): gaps could make a strong difference below the heterogeneous canopies of these forests, soil was extremely heterogeneous with large rocks, modifying soil depth drastically within distances of a few centimetres. The variability between plots was often difficult to interpret and was translated into high plot effects in our model. These could be due to many factors difficult to record: past management history, soil conditions and complex climate effects poorly represented in the interpolated climate data to name a few.
4.2 Sapling height increments
The maximum annual height increment reached by a sapling was primarily determined by its height, canopy openness of the plot and maximum summer temperature (for spruce and beech).
The height increment of a sapling was limited by its height, with taller saplings showing higher increments. In comparison with studies like Ligot et al. (2013) carried out in forests in the Belgian Ardennes (40 cm/year increment for 250-cm-high beech sapling), sapling height increments in our study were relatively modest (15 to 25 cm/year for a 700-cm-high beech sapling), but comparable to other studies such as Dobrovolny (2016) (16 cm for a 200-cm-high beech sapling) carried out in mountain forests.
When plotted against canopy openness (O), the shape of height increment curves for saplings of different heights were similar for the three species (Fig. 4). In particular at low O (<10%), the three species presented very low growth, with beech growth slightly above fir and spruce. This finding was in contradiction with those of Stancioiu and O’Hara (2006) who reported out-competition of spruce by beech and fir growths, but in agreement with Ligot et al. (2020), who observed comparable height growths for spruce and fir. Fir is known to be more shade tolerant than spruce (Diaci et al. 2020; Klopcic et al. 2012) and is expected to have a low growth rate in shade with a low mortality (Kobe et al. 1995). However, more recent work showed that sapling size (ontogeny) must also be considered in that tolerance to shade (e.g. Niinemets (2006)). Messier et al. (1999) defined a critical size for a given light level above which the sapling do not survive. This critical size is higher for shade-tolerant species. It could also explain why small saplings can easily be found under low canopy openness and that higher saplings are only found under higher canopy openness (Dreyer et al. 2005).
However, height and canopy openness were not the only factors affecting height increment. Being sensitive to summer temperatures (Schurman et al. 2019), spruce grew slower in sites with higher maximum summer temperatures. The growth of beech was also inversely proportional to maximum summer temperatures, as also shown in adult trees by Seynave et al. (2008), which is probably due to its sensitivity to higher temperatures and droughts (Gessler et al. 2006; Maes et al. 2018). We could not see any direct climate effect on fir, possibly due to multiple interacting effects that could not be caught in our models. There is also a possibility that high browsing rates, and hence a high proportion of saplings with zero increments (which were not included in the model), led to an inconclusive effect of any environmental variable on fir sapling increments. Even though spruce and beech have different niches, and occur more frequently in different altitudes, it is crucial to note that summer temperatures affect the growth of regeneration of both species. This trend seems rather general and could potentially affect fir as well (Vitasse et al. 2019).
4.3 Browsing
A significant number of saplings (18.6%) were browsed by wild ungulates such as deer, roe deer and chamois, either at the terminal shoot or at peripheral branches, though the actual impact of these animals on sapling density and growth is difficult to evaluate. However, it does reduce or stunt the growth of a significant number of saplings by chewing off the terminal buds that are sometimes even fatal for small saplings (picture of a representative sapling shown in Fig. 6). Very often, the saplings are also browsed very frequently while they are very young, dying or disappearing even before they are counted for studies like ours. This alters the regeneration composition altogether and could result in loss of data needed to understand the process. This remains a challenge in regeneration studies attempting to understand the effect of browsing.

An old fir sapling, completely browsed every year by animals, thus losing any chance to grow
Establishing a direct relation of ungulate browsing with sapling densities or height increments was hence a difficult task, and we did not succeed to include it in the above models. However, the extent of damage by browsing on the field prompted us to explore browsing rates, which turned out to be concerning. Among the three species that we observed, there was a clear and strong preference of browsers towards fir, followed by beech (comparable to fir), and spruce was seldom touched. The affinity of various ungulates to fir is well studied and documented (Bernard et al. 2017; Heuze et al. 2005; Häsler and Senn 2012; Nagel et al. 2015), with an alarming reduction in fir regeneration in areas of high ungulate population. Beech, however, is not commonly known as one of the popular choices of these animals. This indicates that the browsing pressure was quite high in our field sites, and ungulates were forced to eat beech as there was no fir available anymore. It also highlights once again, the high vulnerability of fir, and possibly beech in the future, to browsing by ungulates.
4.4 Regeneration dynamics and management implications
Saplings of all the three species can grow faster as they grow higher, but their mortality also increases, leading to very few of them reaching a tree recruitment stage. It is not difficult to conclude that these processes could lead to low tree recruitment, and that is exactly what we observed. Only 34 plots out of 152 sampled had trees between the DBH of 7.5–8 cm (assumption is that they have been recruited in the last 5 years), with 5 spruce trees, 45 fir trees and 34 beech trees. Many of them looked evidently old, or unhealthy, thus unlikely that the reported number will survive or grow and contribute to overstory canopy or seed rain. This confirms the concerns of the foresters about limited tree recruits.
It has been well established that the challenge of climate change is already affecting growth and survival of regeneration in forests. The most studied effect is probably that of temperature, as shown in studies all over the world (Schurman et al. 2019). Higher temperatures affect survival of plants and trees, causing species moving to higher elevations to find optimal temperatures (Munier et al. 2010; Tingstad et al. 2015; Benavides et al. 2016). However, the direct effect of temperature is often not so obvious, because temperatures are also involved in potential evapotranspiration. We observed a strong plant response to temperature, as well as variables that change water availability to the plant. Increasing temperature and decreasing precipitation is leading to increased frequency and intensity of droughts, and spring and summer droughts, in the growing season of plants could seriously jeopardise regeneration. Additionally, decreasing snow cover could affect water balance. This could be an indicator that mountains are transitioning from temperature to moisture limitation as discussed by Schurman et al. (2019), making predictions even more challenging as this will increase the weight of forest dynamics on local drivers.
All three species are seen to grow better in canopy openness of more than 10% in the plot, which could be an important management implication, to create big enough gaps to allow saplings to grow fast. At present in our sampling, half sites had canopy openness lower than 9.4% (median). This has also been studied and reported by others such as Diaci et al. (2020). Reducing BA will also reduce water losses by tree transpiration and could lead to a better water balance, favourable to regeneration (e.g. Bréda et al. (1995); Aussenac (2000)).
Our model predictions point out that among spruce, fir and beech, fir saplings have the highest mortality and lowest annual height increments. Added to that, the ungulate browsing is the highest in fir. This could create an advantage for spruce and beech regeneration, thus possibly starting a change in species composition of the stands (Dobrowolska and Bolibok 2019; Häsler and Senn 2012; Bernard et al. 2017; Heuze et al. 2005; Diaci 2002; Senn and Suter 2003; Thrippleton et al. 2017; Weisberg et al. 2005; Cretaz and Kelty 2002). However, spruce and beech are also more sensitive to climatic changes, specially limited by high temperatures and water availability. This could also reduce their chances of better survival. In spite of all this information, in our dataset, and in spite of being highly browsed, the highest observed tree recruits in the last few years (7.5cm < tree DBH <8cm) belong to fir. Tinner et al. (2013) has also specified a much larger range of fir in the past. This could indicate a lower browsing pressure, and more favourable climatic conditions for fir in the past. However, when comparing it with the other two species, it could also be either that fir is more resilient than spruce to the factors that hinder its growth and spread, or is simply a lag in response by fir, or a lag in detection of this change by us.
4.5 Limitations and the way forward
One of the limitations in this study was the uncertainty about the reliability of measured and obtained data. In particular, due to the presence of large rocks, or heterogeneous soil conditions within a plot, soil variables could have been misled. The proxy of canopy openness did not always correspond well to the light available for saplings (Comeau et al. 2006), often due to the presence of intermediate layers of canopy formed by tall understory vegetation, or sometimes because of a wider-than-expected spread of the overstory canopy. Thus, we see in our data, a possibility of low basal areas, and low light levels occurring in the same plot.
The explored variables related to climate were all modelled, as it was unrealistic to directly measure them on-site for such a large area. Instead, we used the platform digitalis to simulate long-term climate on a 50-m spatial grid. This can lead to imprecisions in the climate really prevailing in the different considered sites, in particular in mountainous areas with strong topological effects (Piedallu et. al. 2021 (submitted)). But most importantly the simulated variables give the general climate above tree canopy and not the one experienced by the saplings in the understory, that is to say the micro-climate. Indeed, micro-climate at the soil level can be very different from the climate depending on stand density and structure (Aussenac 2000). Of course, the first affected variable is radiation we approximated by canopy openness, with some concerns we already debated above. Linked to attenuated radiation, air temperatures are buffered by the tree canopy, with lower maximum and higher minimum (Gaudio et al. 2017). Tree canopy also intercepts precipitation resulting in less water available in the understory than predicted by incident climate variables and affecting the stand water balance (Barbier et al. 2009). Therefore, the micro-climate experienced by the sapling has opposite components, in one hand a buffered air temperature limiting extreme events such as frost and heat, and also lowering vapour pressure deficit and PET, and so sapling evapotranspiration and in the other hand a periodic soil water deficit in the upper soil horizons linked to rainfall interception by trees. At the difference of adult trees, as sapling roots cannot reach the deeper soil horizons where water can be still present in drought periods, they are subjected to periodic water constrains not monitored in the simulated general climate variables. All these considerations could explain why we had difficulties to have strong effects of other climate variables on sapling growth and density. Such a lag between the general increase in temperature linked to climate change and the effective shift in understory plant composition (including tree saplings) has already be reported (e.g. Dietz et al. (2020)). We also highlight the necessity to better monitor and understand the actual water regime experienced by the saplings as suggested by the high number of water-related variables having a significant effect in our models.
The variability in field studies such as this is so high that it questions the reliability of the “space for time” assumption if we want to use our results to predict changes in time. Monitoring of sites would be necessary to have more accurate results, but the very low fluxes of regeneration densities as well as tree recruitment require to do the monitoring on large scales or long durations.
5 Conclusions
To our knowledge, this is one of the first studies monitoring the regeneration state of spruce-fir-beech on such a large scale. It gives insights of the likely evolution of forest dynamics and composition in the considered area, with a slightly pessimistic prediction for some areas and species. The three species may be at risk because they respond negatively to factors such as low light conditions, browsing, high temperatures and drought. Spruce and beech suggest a possibility of higher vulnerability in instances of frequent and intense summer heat and drought situations. The strong preference of ungulates for fir puts it in a high risk in case of a rapid increase in ungulate browsing. We could therefore expect a diminution of adult tree recruitment and adult stand density of either species depending on local water and browsing conditions. This real diminution of adult density could lead to an increase of light under canopy beneficial to saplings, which could slow the changes. We could have a kind of resilience mechanism here that could prevent species exclusion. Smaller density would also mean smaller competition between adult trees and the total production could remain moreover the same for a while. However, we do not know how far such a mechanism could compensate the effect of recruitment diminution, with a potential risk of changing forest structures, with migration of species at one point.
Availability of data and materials
The datasets generated during and/or analysed during the current study are available on the INRAE data portal www.data.inrae.frwith the following reference: https://doi.org/10.15454/J0NVIK.
Code availability
The code for all analyses was written in R software version 3.6.0 and version 4.0.2 (R Core Team (2017)). The function ``nlme'' from the R package ``nlme'' (Pinheiro et al. (2021)) was used to run models. All figures were produced using the package ggplot2 (Wickham (2009)). The custom code will be made available by the authors on reasonable request.
References
Allen, CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EHT, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259(4):660–684. https://doi.org/10.1016/j.foreco.2009.09.001.
Aussenac, G (2000) Interactions between forest stands and microclimate: ecophysiological aspects and consequences for silviculture. Ann For Sci 57(3):287–301. https://doi.org/10.1051/forest:2000119.
Balandier, P, Collet C, Miller JH, Reynolds PE, Zedaker SM (2006) Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. For Int J For Res 79(1):3–27. https://doi.org/10.1093/forestry/cpi056.
Barbier, S, Balandier P, Gosselin F (2009) Influence of several tree traits on rainfall partitioning in temperate and boreal forests: a review. Ann For Sci 66(6):602–602. https://doi.org/10.1051/forest/2009041.
Baudry, O, Charmetant C, Collet C, Ponette Q (2013) Estimating light climate in forest with the convex densiometer: operator effect, geometry and relation to diffuse light. Eur J For Res 133(1):101–110. https://doi.org/10.1007/s10342-013-0746-6.
Benavides, R, Escudero A, Coll L, Ferrandis P, Ogaya R, Gouriveau F, Peñuelas J, Valladares F (2016) Recruitment patterns of four tree species along elevation gradients in Mediterranean mountains: not only climate matters. Forest Ecol Manag 360:287–296. https://doi.org/10.1016/j.foreco.2015.10.043.
Bernard, M, Boulanger V, Dupouey J-L, Laurent L, Montpied P, Morin X, Picard J-F, Saïd S (2017) Deer browsing promotes Norway spruce at the expense of silver fir in the forest regeneration phase. For Ecol Manag 400:269–277. https://doi.org/10.1016/j.foreco.2017.05.040.
Bodin, P, Wiman BLB (2007) The usefulness of stability concepts in forest management when coping with increasing climate uncertainties. For Ecol Manag 242(2-3):541–552. https://doi.org/10.1016/j.foreco.2007.01.066.
Bottero, A, Forrester DI, Cailleret M, Kohnle U, Gessler A, Michel D, Bose AK, Bauhus J, Bugmann H, Cuntz M, Gillerot L, Hanewinkel M, Lévesque M, Ryder J, Sainte-Marie J, Schwarz J, Yousefpour R, Zamora-Pereira JC, Rigling A (2021) Growth resistance and resilience of mixed silver fir and Norway spruce forests in central Europe: contrasting responses to mild and severe droughts. Glob Chang Biol 27(18):4403–4419. https://doi.org/10.1111/gcb.15737.
Bréda, N, Granier A, Aussenac G (1995) Effects of thinning on soil and tree water relations, transpiration and growth in an oak forest (Quercus petraea (Matt.) Liebl). Tree Physiol 15(5):295–306. https://doi.org/10.1093/treephys/15.5.295.
Brus, DJ, Hengeveld GM, Walvoort DJJ, Goedhart PW, Heidema AH, Nabuurs GJ, Gunia K (2011) Statistical mapping of tree species over Europe. Eur J For Res 131(1):145–157. https://doi.org/10.1007/s10342-011-0513-5.
Cailleret, M, Nourtier M, Amm A, Durand-Gillmann M, Davi H (2013) Drought-induced decline and mortality of silver fir differ among three sites in Southern France. Ann For Sci 71(6):643–657. https://doi.org/10.1007/s13595-013-0265-0.
Carnet, B, Claessens H, Courbaud B (2018) Estimation du potentiel du calcul bayésien approché pour la modélisation de la dynamique des peuplements forestiers : Cas de la régénération naturelle des futaies irrégulières montagnardes (Alpes françaises). (Unpublished master’s thesis). Université de Liège, Liège, Belgique. https://matheo.uliege.be/handle/2268.2/5124.
Caron, MM, Zellweger F, Verheyen K, Baeten L, Hédl R, Bernhardt-Römermann M, Berki I, Brunet J, Decocq G, Díaz S, Dirnböck T, Durak T, Heinken T, Jaroszewicz B, Kopecký M, Lenoir J, Macek M, Malicki M, Máliš F, Nagel TA, Perring MP, Petřík P, Reczyńska K, Pielech R, Schmidt W, Świerkosz K, Teleki B, Wulf M, Frenne PD (2021) Thermal differences between juveniles and adults increased over time in European forest trees. J Ecol 109(11):3944–3957. https://doi.org/10.1111/1365-2745.13773.
Cienciala, E, Lindroth A, Čermák J, Hällgren J-E, Kučera J (1992) Assessment of transpiration estimates for Picea abies trees during a growing season. Trees 6(3):121–127. https://doi.org/10.1007/bf00202427.
Collet, C, Chenost C (2006) Using competition and light estimates to predict diameter and height growth of naturally regenerated beech seedlings growing under changing canopy conditions. Forestry 79(5):489–502. https://doi.org/10.1093/forestry/cpl033.
Comeau, P, Heineman J, Newsome T (2006) Evaluation of relationships between understory light and aspen basal area in the British Columbia central interior. For Ecol Manag 226(1-3):80–87. https://doi.org/10.1016/j.foreco.2005.12.060.
Cretaz, AL, Kelty MJ (2002) Development of tree regeneration in fern-dominated forest understories after reduction of deer browsing. Restor Ecol 10(2):416–426. https://doi.org/10.1046/j.1526-100x.2002.02037.x.
Defossez, E, Courbaud B, Lasbouygues O, Schiffers K, Kunstler G (2015) Are variations of direct and indirect plant interactions along a climatic gradient dependent on species’ strategies? An experiment on tree seedlings. Oikos 125(5):708–717. https://doi.org/10.1111/oik.02331.
Dey, DC, Knapp BO, Battaglia MA, Deal RL, Hart JL, O’Hara KL, Schweitzer CJ, Schuler TM (2018) Barriers to natural regeneration in temperate forests across the USA. New Forest 50(1):11–40. https://doi.org/10.1007/s11056-018-09694-6.
Diaci, J (2002) Regeneration dynamics in a Norway spruce plantation on a silver fir-beech forest site in the Slovenian alps. For Ecol Manag 161(1-3):27–38. https://doi.org/10.1016/s0378-1127(01)00492-3.
Diaci, J, Rozman J, Rozman A (2020) Regeneration gap and microsite niche partitioning in a high alpine forest: are Norway spruce seedlings more drought-tolerant than beech seedlings?For Ecol Manag 455:117688. https://doi.org/10.1016/j.foreco.2019.117688.
Dietz, L, Collet C, Dupouey J-L, Lacombe E, Laurent L, Gégout J-C (2020) Windstorm-induced canopy openings accelerate temperate forest adaptation to global warming. Glob Ecol Biogeogr 29(11):2067–2077. https://doi.org/10.1111/geb.13177.
Dobrovolny, L (2016) Density and spatial distribution of beech (Fagus sylvatica L,) regeneration in Norway spruce (Picea abies (L.) Karsten) stands in the central part of the Czech Republic. iForest Biogeosci For 9(4):666–672. https://doi.org/10.3832/ifor1581-008.
Dobrowolska, D, Bolibok L (2019) Is climate the key factor limiting the natural regeneration of silver fir beyond the northeastern border of its distribution range?For Ecol Manag 439:105–121. https://doi.org/10.1016/j.foreco.2019.02.040.
Dreyer, E, Collet C, Montpied P, Sinoquet H (2005) Caractérisation de la tolérance à l’ombrage de jeunes semis de hêtre et comparaison avec les essences associées. Rev For Fr 2:175. https://doi.org/10.4267/2042/5034.
Dyderski, MK, Gazda A, Hachułka M, Horodecki P, Kałucka IL, Kamczyc J, Malicki M, Pielech R, Smoczyk M, Skorupski M, Wierzcholska S, Jagodziński AM (2018) Impacts of soil conditions and light availability on natural regeneration of Norway spruce Picea abies (L.) H. Karst. in low-elevation mountain forests. Ann For Sci 75(4). https://doi.org/10.1007/s13595-018-0775-x.
Gaudio, N, Gendre X, Saudreau M, Seigner V, Balandier P (2017) Impact of tree canopy on thermal and radiative microclimates in a mixed temperate forest: a new statistical method to analyse hourly temporal dynamics. Agric Forest Meteorol 237-238:71–79. https://doi.org/10.1016/j.agrformet.2017.02.010.
Gessler, A, Keitel C, Kreuzwieser J, Matyssek R, Seiler W, Rennenberg H (2006) Potential risks for European beech (Fagus sylvatica L,) in a changing climate. Trees 21(1):1–11. https://doi.org/10.1007/s00468-006-0107-x.
Gray, SB, Brady SM (2016) Plant developmental responses to climate change. Dev Biol 419(1):64–77. https://doi.org/10.1016/j.ydbio.2016.07.023.
Häsler, H, Senn J (2012) Ungulate browsing on European silver fir (Abies alba): the role of occasions, food shortage and diet preferences. Wildl Biol 18(1):67–74. https://doi.org/10.2981/09-013.
Heuze, P, Schnitzler A, Klein F (2005) Is browsing the major factor of silver fir decline in the Vosges mountains of France?For Ecol Manag 217(2-3):219–228. https://doi.org/10.1016/j.foreco.2005.06.003.
Klopcic, M, Poljanec A, Boncina A (2012) Modelling natural recruitment of European beech (Fagus sylvatica L). Forest Ecol Manag 284:142–151. https://doi.org/10.1016/j.foreco.2012.07.049.
Kobe, RK, Pacala SW, Silander JA, Canham CD (1995) Juvenile tree survivorship as a component of shade tolerance. Ecol Appl 5(2):517–532. https://doi.org/10.2307/1942040.
Kräuchi, N, Brang P, Schönenberger W (2000) Forests of mountainous regions: gaps in knowledge and research needs. For Ecol Manag 132(1):73–82. https://doi.org/10.1016/s0378-1127(00)00382-0.
Kunstler, G, Guyennon A, Ratcliffe S, Rüger N, Ruiz-Benito P, Childs DZ, Dahlgren J, Lehtonen A, Thuiller W, Wirth C, Zavala MA, Salguero-Gomez R (2020) Demographic performance of European tree species at their hot and cold climatic edges. J Ecol 109(2):1041–1054. https://doi.org/10.1111/1365-2745.13533.
Laurent, L, Mårell A, Korboulewsky N, Saïd S, Balandier P (2017) How does disturbance affect the intensity and importance of plant competition along resource gradients?. Forest Ecol Manag 391:239–245. https://doi.org/10.1016/j.foreco.2017.02.003.
Ligot, G, Balandier P, Courbaud B, Jonard M, Kneeshaw D, Claessens H (2014) Managing understory light to maintain a mixture of species with different shade tolerance. Forest Ecol Manag 327:189–200. https://doi.org/10.1016/j.foreco.2014.05.010.
Ligot, G, Balandier P, Fayolle A, Lejeune P, Claessens H (2013) Height competition between Quercus petraea and Fagus sylvatica natural regeneration in mixed and uneven-aged stands. Forest Ecol Manag 304:391–398. https://doi.org/10.1016/j.foreco.2013.05.050.
Ligot, G, Balandier P, Schmitz S, Claessens H (2020) Transforming even-aged coniferous stands to multi-aged stands: an opportunity to increase tree species diversity?For Int J For Res 93(5):616–629. https://doi.org/10.1093/forestry/cpaa004.
Lines, ER, Zavala MA, Ruiz-Benito P, Coomes DA (2019) Capturing juvenile tree dynamics from count data using approximate Bayesian computation. Ecography 43(3):406–418. https://doi.org/10.1111/ecog.04824.
Lochhead, KD, Comeau PG (2012) Relationships between forest structure, understorey light and regeneration in complex douglas-fir dominated stands in south-eastern British Columbia. Forest Ecol Manag 284:12–22. https://doi.org/10.1016/j.foreco.2012.07.029.
Löf, M, Bolte A, Welander NT (2005) Interacting effects of irradiance and water stress on dry weight and biomass partitioning in Fagus sylvatica seedlings. Scand J For Res 20(4):322–328. https://doi.org/10.1080/02827580500201593.
Löf, M, Madsen P, Metslaid M, Witzell J, Jacobs DF (2019) Restoring forests: regeneration and ecosystem function for the future. New Forest 50(2):139–151. https://doi.org/10.1007/s11056-019-09713-0.
Löf, M, Welander NT (2000) Carry-over effects on growth and transpiration in Fagus sylvatica seedlings after drought at various stages of development. Can J For Res 30(3):468–475. https://doi.org/10.1139/x99-232.
Lombaerde, ED, Blondeel H, Baeten L, Landuyt D, Perring MP, Depauw L, Maes SL, Wang B, Verheyen K (2020) Light, temperature and understorey cover predominantly affect early life stages of tree seedlings in a multifactorial mesocosm experiment. For Ecol Manag 461:117907. https://doi.org/10.1016/j.foreco.2020.117907.
Madsen, P, Larsen JB (1997) Natural regeneration of beech (Fagus sylvatica L,) with respect to canopy density, soil moisture and soil carbon content. For Ecol Manag 97(2):95–105. https://doi.org/10.1016/s0378-1127(97)00091-1.
Maes, SL, Perring MP, Vanhellemont M, Depauw L, den Bulcke JV, Brūmelis G, Brunet J, Decocq G, den Ouden J, Härdtle W, Hédl R, Heinken T, Heinrichs S, Jaroszewicz B, Kopecký M, Máliš F, Wulf M, Verheyen K (2018) Environmental drivers interactively affect individual tree growth across temperate European forests. Glob Change Biol 25(1):201–217. https://doi.org/10.1111/gcb.14493.
Manso, R, Morneau F, Ningre F, Fortin M (2015) Incorporating stochasticity from extreme climatic events and multi-species competition relationships into single-tree mortality models. For Ecol Manag 354:243–253. https://doi.org/10.1016/j.foreco.2015.06.008.
Messier, C, Doucet R, Ruel J-C, Claveau Y, Kelly C, Lechowicz MJ (1999) Functional ecology of advance regeneration in relation to light in boreal forests. Can J For Res 29(6):812–823. https://doi.org/10.1139/x99-070.
Mitchell, K (2015) Quantitative analysis by the point-centered quarter method. http://arxiv.org/abs/1010.3303.
Moser, B, Walthert L, Metslaid M, Wasem U, Wohlgemuth T (2016) Spring water deficit and soil conditions matter more than seed origin and summer drought for the establishment of temperate conifers. Oecologia 183(2):519–530. https://doi.org/10.1007/s00442-016-3766-3.
Munier, A, Hermanutz L, Jacobs JD, Lewis K (2010) The interacting effects of temperature, ground disturbance, and herbivory on seedling establishment: implications for treeline advance with climate warming. Plant Ecol 210(1):19–30. https://doi.org/10.1007/s11258-010-9724-y.
Nagel, TA, Diaci J, Jerina K, Kobal M, Rozenbergar D (2015) Simultaneous influence of canopy decline and deer herbivory on regeneration in a conifer–broadleaf forest. Can J For Res 45(3):266–275. https://doi.org/10.1139/cjfr-2014-0249.
Newcombe, RG (1998) Interval estimation for the difference between independent proportions: comparison of eleven methods. Stat Med 17(8):873–890. https://doi.org/10.1002/(sici)1097-0258(19980430)17:8<873::aid-sim779>3.0.co;2-i.
Niinemets, U (2006) The controversy over traits conferring shade-tolerance in trees: ontogenetic changes revisited. J Ecol 94(2):464–470. https://doi.org/10.1111/j.1365-2745.2006.01093.x.
Orman, O, Wrzesiński P, Dobrowolska D, Szewczyk J (2021) Regeneration growth and crown architecture of European beech and silver fir depend on gap characteristics and light gradient in the mixed montane old-growth stands. Forest Ecol Manag 482:118866. https://doi.org/10.1016/j.foreco.2020.118866.
Paluch, J, Bartkowicz L, Moser WK (2019) Interspecific effects between overstorey and regeneration in small-scale mixtures of three late-successional species in the Western Carpathians (southern Poland). Eur J For Res 138(5):889–905. https://doi.org/10.1007/s10342-019-01209-y.
Piedallu, C, Gégout J-C (2007) Multiscale computation of solar radiation for predictive vegetation modelling. Ann For Sci 64(8):899–909. https://doi.org/10.1051/forest:2007072.
Piedallu, C, Gégout J-C, Lebourgeois F, Seynave I (2016) Soil aeration, water deficit, nitrogen availability, acidity and temperature all contribute to shaping tree species distribution in temperate forests. J Veg Sci 27(2):387–399. https://doi.org/10.1111/jvs.12370.
Piedallu, C, Gégout J-C, Perez V, Lebourgeois F (2013) Soil water balance performs better than climatic water variables in tree species distribution modelling. Glob Ecol Biogeogr 22(4):470–482. https://doi.org/10.1111/geb.12012.
Pinheiro, J, Bates D, DebRoy S, Sarkar D, R Core Team (2021) nlme: linear and nonlinear mixed effects models. https://CRAN.R-project.org/package=nlme.
Pröll, G, Hietz P, Delaney CM, Katzensteiner K (2015) Substrate influences ecophysiological performance of tree seedlings. Tree Physiol 36(1):39–53. https://doi.org/10.1093/treephys/tpv104.
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/.
Schall, P, Lödige C, Beck M, Ammer C (2012) Biomass allocation to roots and shoots is more sensitive to shade and drought in European beech than in Norway spruce seedlings. For Ecol Manag 266:246–253. https://doi.org/10.1016/j.foreco.2011.11.017.
Schurman, JS, Babst F, Björklund J, Rydval M, Bače R, Čada V, Janda P, Mikolas M, Saulnier M, Trotsiuk V, Svoboda M (2019) The climatic drivers of primary Picea forest growth along the carpathian arc are changing under rising temperatures. Glob Chang Biol 25(9):3136–3150. https://doi.org/10.1111/gcb.14721.
Senn, J, Suter W (2003) Ungulate browsing on silver fir (Abies alba) in the Swiss alps: beliefs in search of supporting data. For Ecol Manag 181(1-2):151–164. https://doi.org/10.1016/s0378-1127(03)00129-4.
Seynave, I, Gégout J-C, Hervé J-C, Dhôte J-F (2008) Is the spatial distribution of European beech (Fagus sylvatica L,) limited by its potential height growth?J Biogeogr 35(10):1851–1862. https://doi.org/10.1111/j.1365-2699.2008.01930.x.
Silva, DE, Mazzella PR, Legay M, Corcket E, Dupouey JL (2012) Does natural regeneration determine the limit of European beech distribution under climatic stress?. For Ecol Manag 266:263–272. https://doi.org/10.1016/j.foreco.2011.11.031.
Simon, A, Katzensteiner K, Gratzer G (2019) Drivers of forest regeneration patterns in drought prone mixed-species forests in the Northern Calcareous Alps. For Ecol Manag 453:117589. https://doi.org/10.1016/j.foreco.2019.117589.
Stancioiu, PT, O’Hara KL (2006) Regeneration growth in different light environments of mixed species, multi-aged, mountainous forests of Romania. Eur J For Res 125(2):151–162. https://doi.org/10.1007/s10342-005-0069-3.
Thornthwaite, CW (1955) The water balance. Climatology.
Thrippleton, T, Bugmann H, Snell RS (2017) Herbaceous competition and browsing may induce arrested succession in central European forests. J Ecol 106(3):1120–1132. https://doi.org/10.1111/1365-2745.12889.
Tingstad, L, Olsen SL, Klanderud K, Vandvik V, Ohlson M (2015) Temperature, precipitation and biotic interactions as determinants of tree seedling recruitment across the tree line ecotone. Oecologia 179(2):599–608. https://doi.org/10.1007/s00442-015-3360-0.
Tinner, W, Colombaroli D, Heiri O, Henne PD, Steinacher M, Untenecker J, Vescovi E, Allen JRM, Carraro G, Conedera M, Joos F, Lotter AF, Luterbacher J, Samartin S, Valsecchi V (2013) The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol Monogr 83(4):419–439. https://doi.org/10.1890/12-2231.1.
Toïgo, M, Vallet P, Perot T, Bontemps J-D, Piedallu C, Courbaud B (2015) Overyielding in mixed forests decreases with site productivity. J Eclect 103(2):502–512. https://doi.org/10.1111/1365-2745.12353.
Turc, L (1961) Estimation of irrigation water requirements, potential evapotranspiration: a simple climatic formula evolved up to date. Ann Agron 12(1):13–49.
Unkule, M, Courbaud B, Balandier P, Mermin E, Barier R, Ruth T (2021) Regeneration and related variables data in spruce-fir-beech mixed uneven aged forests in the French Alps. [dataset]. Portail Data INRAE, V1. https://doi.org/10.15454/J0NVIK.
Vitali, V, Büntgen U, Bauhus J (2017) Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob Chang Biol 23(12):5108–5119. https://doi.org/10.1111/gcb.13774.
Vitasse, Y, Bottero A, Rebetez M, Conedera M, Augustin S, Brang P, Tinner W (2019) What is the potential of silver fir to thrive under warmer and drier climate?. Eur J For Res 138(4):547–560. https://doi.org/10.1007/s10342-019-01192-4.
Warde, W, Petranka JW (1981) A correction factor table for missing point-center quarter data. Ecology 62(2):491–494. https://doi.org/10.2307/1936723.
Weisberg, PJ, Bonavia F, Bugmann H (2005) Modeling the interacting effects of browsing and shading on mountain forest tree regeneration (Picea abies). Ecol Model 185(2-4):213–230. https://doi.org/10.1016/j.ecolmodel.2004.12.004.
Wickham, H (2009) Ggplot2: elegant graphics for data analysis. Springer. http://ggplot2.org.
Zang, C, Hartl-Meier C, Dittmar C, Rothe A, Menzel A (2014) Patterns of drought tolerance in major European temperate forest trees: climatic drivers and levels of variability. Glob Chang Biol 20(12):3767–3779. https://doi.org/10.1111/gcb.12637.
Acknowledgements
We thank the French National Forests Office (ONF project GESTFOR) and INRAE who co-funded this project. The help of Eric Mermin, Rachel Barrier, Ruth Tanguy and Yann Dumas was invaluable in field data collection and laboratory analyses.
Funding
This study was co-funded by French National Forests Office (ONF project GESTFOR) and INRAE, LESSEM.
Author information
Authors and Affiliations
Contributions
BC, MU and PB conceptualised and designed the study. MU carried out the data collection with help from BC and PB. BC wrote the primary R codes for analyses, and the analyses were carried forward by MU. All the climate and soil satellite data was obtained by CP. MU wrote the manuscript with inputs from BC, PB and CP. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
The authors of this manuscript give their consent for publication of the manuscript on an open access basis. We understand that it will be freely available on the Internet and be seen by the public.
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling Editor: Andreas Bolte
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Unkule, M., Piedallu, C., Balandier, P. et al. Climate and ungulate browsing impair regeneration dynamics in spruce-fir-beech forests in the French Alps. Annals of Forest Science 79, 11 (2022). https://doi.org/10.1186/s13595-022-01126-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13595-022-01126-y